
Celleadhæsionsproteiner
cellecelleadhæsion er en selektiv proces, således at celler kun klæber til andre celler af specifikke typer. Denne selektivitet blev først demonstreret i klassiske studier af embryonudvikling, som viste, at celler fra et væv (f.eks. lever) specifikt klæber til celler i det samme væv snarere end til celler i et andet væv (f. eks. hjerne). En sådan selektiv cellecelleadhæsion medieres af transmembranproteiner kaldet celleadhæsionsmolekyler, som kan opdeles i fire hovedgrupper: selectinerne, integrinerne, immunoglobulinet (ig) superfamilien (så navngivet, fordi de indeholder strukturelle domæner svarende til immunoglobuliner) og cadherinerne (tabel 12.3). Celleadhæsion medieret af selectiner, integriner og cadheriner kræver Ca2+ eller Mg2+, så mange klæbende interaktioner mellem celler er Ca2+- eller Mg2+-afhængige.
tabel 12.3
Celleadhæsionsmolekyler.
selectinerne formidler forbigående interaktioner mellem leukocytter og endotelceller eller blodplader. Der er tre medlemmer af selectinfamilien: L-selectin, som udtrykkes på leukocytter; E-selectin, som udtrykkes på endotelceller; og P-selectin, som udtrykkes på blodplader. Som diskuteret tidligere i dette kapitel genkender selectinerne celleoverfladecarbohydrater (se figur 12.14). En af deres kritiske roller er at indlede interaktionerne mellem leukocytter og endotelceller under migrering af leukocytter fra cirkulationen til steder med vævsbetændelse (figur 12.62). Selectinerne medierer den indledende adhæsion af leukocytter til endotelceller. Dette efterfølges af dannelsen af mere stabile adhæsioner, hvor integriner på overfladen af leukocytter binder til intercellulære adhæsionsmolekyler (ICAMs), som er medlemmer af IG-superfamilien udtrykt på overfladen af endotelceller. De fast fastgjorte leukocytter er derefter i stand til at trænge ind i væggene i kapillærerne og komme ind i det underliggende væv ved at migrere mellem endotelceller.

figur 12.62
adhæsion mellem leukocytter og endotelceller. Leukocytter forlader cirkulationen på steder med vævsbetændelse ved at interagere med endotelcellerne i kapillærvægge. Det første trin i denne interaktion er bindingen af leukocyt selectiner (mere…)
bindingen af ICAMs til integriner er et eksempel på en heterofile interaktion, hvor et adhæsionsmolekyle på overfladen af en celle (f.eks. en ICAM) genkender et andet molekyle på overfladen af en anden celle (f. eks. et integrin). Andre medlemmer af IG superfamilien medierer homofile interaktioner, hvor et adhæsionsmolekyle på overfladen af en celle binder til det samme molekyle på overfladen af en anden celle. En sådan homofil binding fører til selektiv adhæsion mellem celler af samme type. For eksempel er nervecelleadhæsionsmolekyler (N-CAMs) medlemmer af IG-superfamilien udtrykt på nerveceller, og homofil binding mellem N-CAMs bidrager til dannelsen af selektive foreninger mellem nerveceller under udvikling. Der er mere end 100 medlemmer af IG superfamilien, som formidler en række celle-celle interaktioner.
den fjerde gruppe af celleadhæsionsmolekyler, cadherinerne, viser også homofile bindingsspecifikationer. De er ikke kun involveret i selektiv vedhæftning mellem embryonale celler, men er også primært ansvarlige for dannelsen af stabile kryds mellem celler i væv. For eksempel udtrykkes E-cadherin på epithelceller, så homofile interaktioner mellem E-cadheriner fører til selektiv adhæsion af epithelceller til hinanden. Det er bemærkelsesværdigt, at tab af E-cadherin kan føre til udvikling af kræftformer, der opstår fra epitelceller, hvilket illustrerer vigtigheden af Celle-Celle-interaktioner i kontrol af celleadfærd. Forskellige medlemmer af cadherin-familien, såsom N-cadherin (neural cadherin) og P-cadherin (placental cadherin), formidler selektiv vedhæftning af andre celletyper.
omkring tyve forskellige klassiske cadheriner, såsom E-cadherin, er blevet identificeret. Derudover udtrykkes en særskilt underfamilie af cadheriner, kaldet protocadheriner, i centralnervesystemet, hvor de ser ud til at spille en rolle i vedhæftning mellem neuroner ved synapser. Spændende synes forskellige neuroner at udtrykke forskellige protocadheriner, hvilket tyder på, at protocadherinerne kan spille en rolle i etableringen af specifikke forbindelser mellem neuroner. Cirka 50 humane protocadheringener er blevet identificeret og vist sig at være organiseret i tre genklynger. Hver klynge indeholder flere eksoner, der koder for De N-terminale ekstracellulære og transmembrane protocadherin-domæner, men kun et enkelt sæt af tre eksoner, der koder for det C-terminale cytoplasmatiske domæne (figur 12.63). Protocadherin-genklyngerne ser således ud til at bestå af en variabel region, der koder for flere ekstracellulære og transmembrane domæner, knyttet til en konstant region, der koder for et enkelt cytoplasmatisk domæne. Denne organisation af protocadheringener ligner påfaldende den for immunoglobulin-og T-cellereceptorgener (se figur 5.42 og 5.43), hvor flere variable regionseksoner er forbundet med en enkelt konstant regionsekson. I immunoglobulin-og T-cellereceptorgenerne sker dette som et resultat af DNA-omlejringer, der genererer mangfoldighed i immunsystemet. Det skal stadig bestemmes, om de variable og konstante regioner af protocadheriner er forbundet på DNA-eller RNA-niveauet (for eksempel ved alternativ splejsning), og i hvilket omfang omlejringer af protocadheringener kan bidrage til etablering af specifikke synaptiske forbindelser i hjernen.
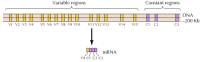
figur 12.63
organisering af protocadherin-genklynger. De humane protocadheringener er organiseret i tre klynger. I den illustrerede klynge er 15 forskellige variable regioner, der koder for ekstracellulære og transmembrane domæner, knyttet til en enkelt konstant region (mere…)
i modsætning til de stabile cellematriceforbindelser, der er diskuteret i det foregående afsnit, er cellecelleinteraktionerne medieret af selectinerne, integrinerne og medlemmerne af IG superfamilien forbigående adhæsioner, hvor cytoskeletterne i tilstødende celler ikke er knyttet til hinanden. Stabile adhæsionskryds, der involverer cytoskeletterne i tilstødende celler, medieres i stedet af cadherinerne. Som omtalt i kapitel 11 er disse cellecelleforbindelser af to typer: adherens-kryds og desmosomer, hvor cadheriner eller beslægtede proteiner (desmogleiner og desmocolliner) er knyttet til henholdsvis actinbundter og mellemfilamenter (figur 12.64). Cadherins rolle i at forbinde cytoskeletterne i tilstødende celler er således analog med integrinernes rolle ved dannelse af stabile kryds mellem celler og den ekstracellulære matrice.

Figur 12.64
stabile cellecelleforbindelser medieret af cadherinerne. Homofile interaktioner mellem cadheriner medierer to typer stabile Celle-Celle adhæsioner. I adherens kryds er cadherinerne knyttet til bundter af actinfilamenter via cateninerne (se figur (mere…)