
Lepší malém měřítku kontinuální kultivace systému
V této studii jsme se ukázat další pokrok a možnosti použití malého rozsahu kontinuální kultivace systému, dříve vyvinut Klein et al. , pro zvýšený stupeň paralelizace a lepší manipulaci a monitorování jednotlivých reaktorů. Klíčové aspekty změn provedených v systému zahrnují zvýšení sady paralelních kultivačních nádob z 8 na 24 reaktorů a snížení pracovního objemu z 10 na 6,5 ml. Současný systém se dále skládá z vlastní-made víčka bydlení čtyři pevné porty používané pro větrání, dodávky médií, vývar odstranění, stejně jako očkování nebo odběru vzorků (Obr. 1). Kromě čtyř portů, optický tyčinkovité DĚLAT sonda se vloží přes víko pro sledování bez narušení proces pěstování, a tímto způsobem se výměna kyslíku fluorescenční senzor místě předchozího nastavení . Vodní lázně, která v předchozím set-up udržuje stálou kultivační teplotu, byl nahrazen na zakázku hliníkové ohřívací blok, který je chráněn pojistkou mikrodestičky míchadlo jednotky. Jako předchozí verze malého rozsahu bioreaktoru systém byl ověřen pomocí štěpení kvasinky Schizosaccharomyces pombe , jsme tady představit lepší pěstování set-up pro S. cerevisiae kultivace.
byly provedeny základní provozní kroky, jakož i úpravy rychlosti ředění výběrem vhodného průměru trubky a rychlosti čerpadla Čerpadla přítoku média, jak bylo popsáno výše . Zde byla hmotnost kapalného obsahu každého bioreaktoru stanovena gravimetricky na konci kultivace, což umožnilo přesný výpočet příslušné rychlosti ředění s 5.1% odchylka. Kultivační vývar a plynná fáze reaktoru byly odstraněny stejným otvorem víka reaktoru pomocí efluxního čerpadla (obr. 1). Pro všechny kultivační experimenty byly použity rychlosti efluxní pumpy 7,5 mL min−1. Rychlost efluxního čerpadla byla daleko vyšší než rychlost napájecího čerpadla a vytvářela mírný podtlak uvnitř kultivační nádoby. Tento tlakový rozdíl vedl k přítoku vzduchu provzdušňovacím portem. Průměrný dosažený koeficient přenosu hmotnosti kyslíku kLa byl 110 h-1, což umožnilo hladiny DO výrazně nad 30% saturací během kultivačního procesu. Hodnota pH nebyla sledována on-line, ani kontrolována během pěstování, jako médium byl a priori upraveny na pH 6,0, což vyústilo v konečné pH 5.5 v pěstování vývar. PH bylo denně měřeno na lince od odtoku reaktorů a po sklizni. PH zůstalo konstantní, jakmile bylo dosaženo ustáleného stavu a odpadní voda z reaktoru vykazovala malou odchylku 0,1 jednotek pH (údaje nejsou uvedeny).
Zkoumání 3-HP produkci v malém měřítku chemostats v různých ředění sazby pod C-omezující podmínky
určit maximální specifické růstové rychlosti (µmax), S. cerevisiae ST938 byla pěstována v dávkovém podmínky použití nadměrné dostupnost živin. Při použití glukózy jako zdroje uhlíku byl µmax 0,265 h-1, výtěžek biomasy byl 24,9 g mol-1 a výtěžek uhlíku 3-HP na glukóze byl 0,6 % C-mol (Tabulka 1). V tomto kultivačním režimu, většina uhlíku byla metabolizována na ethanol a CO2, jako vysoké glykolytické toky v divokém typu s. cerevisiae jsou silně spojeny s alkoholovou fermentací . Kultivace chemostatu jsou řízeny dodávkou substrátu omezujícího růst. V ustáleném stavu, wild-type S. cerevisiae neprodukuje významné množství přetečení metabolitů pod určitou rychlost růstu, označení kritické ředění sazba (Dcrit (h−1)) vzhledem k nedostatku akumulace zdroj uhlíku. Na nebo nad tuto kritickou ředění sazba zdroj uhlíku se hromadí v reaktoru a přetečení metabolismus je aktivován, což vede k výrobě různých vedlejších produktů, jako je etanol, acetát a menší množství organických kyselin . V souladu s tím byly použity kontinuální kultury ke zkoumání vztahu mezi rychlostí růstu a tvorbou produktu 3-HP specifického pro biomasu v S. cerevisiae ST938. Čtyři různé ředění sazby D (h−1): 0.04, 0.09, 0.17 a 0.21 byly vybrány, aby se pod µmax S. cerevisiae ST938 stanovena v batch kultivaci. Byla zkoumána výtěžnost biomasy na glukóze, jakož i výtěžnost 3-HP, specifická rychlost produkce 3-HP a specifická rychlost absorpce substrátu (obr. 2a-d, Tabulka 1). Všechny příslušné parametry pěstování, tj. výnosy a míry s příslušnými směrodatnými odchylkami, jsou shrnuty v tabulce 1.
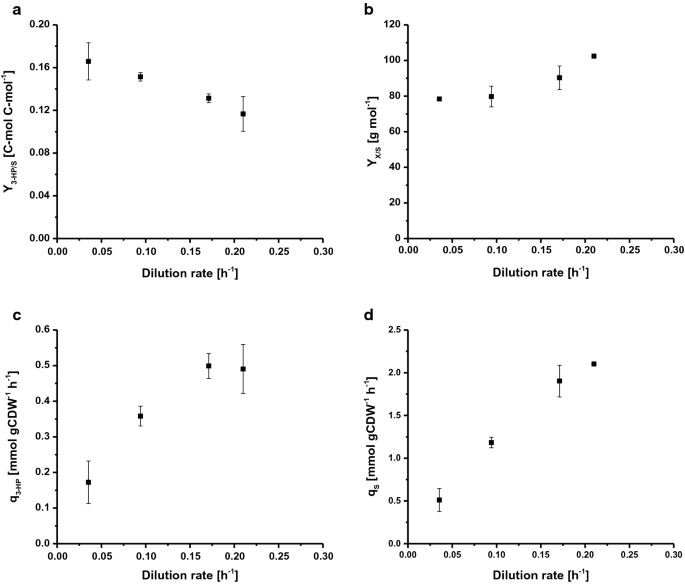
Selected rates and yields for C-limited aerobic chemostat cultivations of S. cerevisiae ST938 a 3-HP yield (C-mol C-mol−1) on glucose, b biomass yield on glucose (g mol−1), c specific 3-HP production rate (mmol gCDW−1 h−1) and d specific glucose uptake rate (mmol gCDW−1 h−1) at different dilution rates for S. cerevisiae ST938. Cultivations were carried out in triplicates at 30 °C and pH 5.5 under C-limited conditions. Chyby odpovídají směrodatné odchylky odvozené z trojím vyhotovení pěstování
kultivaci S. cerevisiae ST938 pod C-omezující podmínky, ukázal přechod z převážně fermentativní metabolismus pozorována v dávkovém režimu respirační metabolismus v chemostats, což se odráží ve vyšší výnosy biomasy, stejně jako v zanedbatelném ethanol a glycerol tvorba (Tabulka 1). Dále menší množství zbytkové glukózy pod 0.Ve vzorcích odebraných z efluxu různých reaktorů bylo zjištěno 1 mM, což ověřilo, že kultury mají být omezeny na glukózu. Uhlí-omezené podmínky našel v ustáleném stavu chemostat kultur zdálo, že podporují tvorbu 3-HP, jako výnosy z tohoto produktu byly 20 – 25-krát vyšší než výnos stanoven v dávce kultury (Tabulka 1). To je s největší pravděpodobností způsobeno účinnější přeměnou uhlíku na energii ve formě ATP během dýchání ve srovnání s fermentací v dávkových kulturách. Zajímavé je, že kultivace s. cerevisiae ST938 pod C-omezující podmínky zjištěno, že s klesající specifické růstové rychlosti konstantní zvýšení v 3-HP uhlíku výnosy by mohlo být pozorováno, s nejvyšší 3-HP výnos 16,6 %C-mol pozorovány v ředění sazba 0,04 h−1 (Obr. 2a). 3-HP výnos biomasy je desetinásobně vyšší než výnos v dávkových kulturách a zvýšil více než dvojnásobně z 0.19 0,43 g gCDW−1 snížením poměru ředění od 0,21 0,04 h−1 (Tabulka 1). Maximální specifické produktivity pro 3-HP přibližně 0.50 mmol gCDW−1 h−1 byl viděn na ředění sazby 0,17 a 0.21 h-1, což je zhruba čtyřnásobně vyšší než měřeno v dávkových kultivacích (obr. 2c, Tabulka 1). Jak se dalo očekávat, specifické vychytávání glukózy sazba (qS) se zvyšuje s vyšší ředění sazby od 0.51 do 2,1 mmol gCDW−1 h−1 (Obr. 2d). Tyto hodnoty jsou až 20krát nižší než maximální specifická rychlost absorpce glukózy 10,7 mmol gCDW-1 h-1 pozorovaná při µmax za podmínek růstu omezeného na glukózu v dávkovém režimu (Tabulka 1). Hodnoty získané pro specifickou rychlost absorpce substrátu a výtěžek biomasy při rychlosti ředění 0.09 h−1 (Tabulka 1) byly ve shodě s údaji z chemostat kultivace s wild-type S. cerevisiae v ředění sazba 0,10 h−1, s uvedením 1.1 a 1,25 mmol gCDW−1 h−1, stejně jako 0.49 g g−1, resp. Bylo zjištěno, že biomasa výnos na substrátu se zvyšuje s rostoucí ředění sazby, pro wild-type S. cerevisiae, nicméně, tento parametr je obecně konstantní níže Dcrit tak dlouho, jako udržovací metabolismus představuje značné uhlíku snížení biomasy výnos při nízkých ředění sazby . V případě s. cerevisiae kmen ST938 se však zdálo, že uhlík, který nebyl zaměřen na tvorbu biomasy, do jisté míry přispívá k tvorbě jak 3-HP, tak vedlejších produktů.
Na závěr rozdíly zažil v kontinuálních kulturách na různých ředění sazby, stejně jako ve srovnání s podobnými set-up s S. cerevisiae wild-type naznačují, že integrované biosyntetické dráhy produkovat 3-HP má obrovský vliv na kvasinky fyziologie, která je pravděpodobně zesílen adaptivní reakce na stres. Konečně, C-omezená kultivace chemostatu při nejnižší rychlosti ředění 0.04 h-1 vedlo k Nejvyššímu výtěžku uhlíku o výkonu 3 HP.
Vyšetřuje dusíku a fosfátu omezení pro zvýšení 3-HP výnosy v malém měřítku chemostat kultur
Po charakterizující S. cerevisiae ST938 pod C-omezující podmínky, pěstování pod dusíku a fosfátu – omezující podmínky byla provedena, jak již dříve bylo prokázáno, že být příznivé pro výrobu různých nativní a non-rodilí metabolitů v různých organismů . Zde byly zkoumány odpovídající kultivace s omezením dusíku (N) A fosforu (P)v malém kontinuálním kultivačním systému (obr. 1) se stanovenou rychlostí ředění 0,04 h-1, což dříve vedlo k nejvyšším výtěžkům uhlíku 3-HP za podmínek omezených C (tabulka 1). Parametry a příslušné směrodatné odchylky od N – A P-omezených kultivací kvasinkového kmene ST938 jsou shrnuty v tabulce 2.
Na N – a P-omezené pěstování podmínky, žádné zbytkové amonný nebo vápenatý byly zjištěny ve vzorcích odebraných z eflux z různých reaktorů, ověření kultur být omezena s ohledem na odpovídající substrát omezení (Tabulka 2). Při omezení N a P však byla zjištěna malá množství zbytkové glukózy v rozmezí 0, 63 A 0, 19 mM (Tabulka 2). Během C-omezení byly zbytkové hladiny glukózy výrazně pod 0,1 mM(Tabulka 2). Kultivace použití N – a P-s omezeními, 3-HP výnos 17.7 a 21,1%, C-mol, respektive zobrazení výrazně vyšší hodnotu, pod P-omezené podmínky ve srovnání s 3-HP uhlíku výnos 16,6 %C-mol vypočteno pro C-omezené pěstování podmínky (Tabulky 1, 2). Podobně specifické produktivity pro 3-HP byla výrazně zvýšena na N – a P-omezená kultivace odhalující hodnoty 0,22 a 0.23 mmol gCDW−1 h−1 ve srovnání se 0.17 mmol gCDW−1 h−1 dosáhl v C-omezený kultur. Výnosy z biomasy o výkonu 3 HP navíc vykazovaly srovnatelné hodnoty 0,43 a 0.44 g gCDW-1 pro C-a n-omezující podmínky, avšak v případě P-omezení byl pozorován zvýšený výtěžek 0,50 g gCDW−1. Vzhledem k celkově nízkým koncentracím kvantifikovaných vedlejších produktů jsou tyto hodnoty zanedbatelné (Tabulka 2). Hodnoty pro konkrétní substrát rychlost příjmu (qS) pro C – a P-omezení byly srovnatelné a mírně zvýšené hodnoty 0,61 mmol gCDW−1 h−1 byla získána v rámci N-omezené podmínky. Biomasy výnosy na substrát ukázal na N – a P-omezené podmínky podobné hodnoty 72,4 a 75.2 g mol−1, resp. Ve srovnání s výtěžností biomasy 78.4 g mol−1 dosažených za podmínek omezených C jsou tyto hodnoty podobné. Naše studie podporuje zdůvodnění, že vyšší výtěžky produktu na substrát byly dosaženy za podmínek omezujících N A P. V důsledku toho, zdá se, že vyšší specifický substrát rychlost příjmu v případě N-omezení v kombinaci s minimální tvorbou vedlejších produktů, nakonec zvýhodněný produkt formace. Za podmínek omezujících P bylo vytvořeno nejmenší množství vedlejších produktů, které by mohly upřednostňovat syntézu produktu. Dále lze spekulovat, že některé metabolické změny mohou vést ke snížení množství uhlíku uvolňovaného jako CO2.
Stručně řečeno, kultivace chemostatu v rámci omezení N a P odhalily zvýšené výnosy 3 HP a specifické míry produkce ve srovnání s podmínkami omezenými C, s omezením P umožňujícím nejvyšší výnosy produktu.
Převoditelnost fyziologické parametry měřené v malém měřítku chemostats fed-batch kultur v 1-L bench-top bioreaktory pod C – a P-omezující podmínky
Protože tato studie dále zaměřena na posouzení srovnatelnosti, přenositelnosti a fyziologické parametry měřené za různých pěstitelských podmínek, pěstování parametry získané z C – a P-omezující podmínky stanovené v chemostat kultury byly převedeny do 1-L míchá bench-top reaktory běží při fed-batch režim. Koncept tohoto přístupu proto spočíval v udržení klíčových podmínek a parametrů konstantních pro chemostatové a krmené dávkové kultury, aby byla zajištěna srovnatelnost. Tyto podmínky se skládá (i) stejná C:P poměr, jak je použito v P-omezení chemostat kultivace, (ii) stejné podmínky procesu, jako je pH a teplota, a (iii) stejné specifické růstové rychlosti, jak je uplatňována v chemostats, dosaženo prostřednictvím exponenciální krmivo profil. Pro optimalizaci titru produktu, parametry s maximálním výtěžkem produktu na substrát z experimentů s chemostatem (D = 0.04 h-1) v rámci C – a p-omezení byly vybrány pro přenos do Fed-dávkového systému. Z důvodu technických omezení v set-up, fed-batch kultivace byly provedeny na tempo růstu ve výši 0,05 h−1, což je mírně vyšší ve srovnání se nastavit ředění sazba 0,04 h−1 v chemostat kultivace. Fed-batch kultivace se skládal z počáteční dávky fáze generovat biomasy následuje exponenciální, živiny omezené krmení fáze kontrolovat tempo růstu. Jen krmení fáze byla považována za relevantní pro posouzení převoditelnosti parametrů získaných z kontinuální kultivace systému, neboť pouze v této fázi, koncentrace substrátu byla kontrolní a tedy omezení růstu mikroorganismů. Kultivační profily krmených šarží s. cerevisiae ST938 za použití podmínek omezujících C A P jsou znázorněny na obr. 3 a odpovídající kultivační parametry s jejich příslušnou směrodatnou odchylkou jsou shrnuty v tabulce 3.
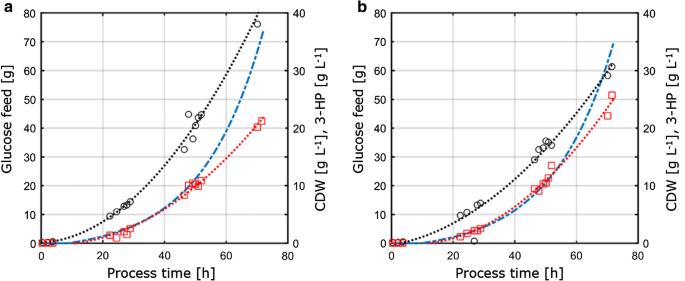
Aerobní fed-batch kultivace v 1-L bench-top fermentory S. cerevisiae ST938 pomocí exponenciální krmení nájezd na D = 0.05 h−1 s omezením substrátu uhlík, nebo b fosforu. Černé kruhy odkazují na biomasu titr, červené čtverce-3-HP titr a modrá přerušovaná čára na absolutní množství glukózy fed reaktorů
biomasy výnosu pozorované ve fed-batch kultur bylo 77.9 a 67.7 g mol−1 pro C – a P-omezení, respektive. Biomasy výnos pro C-omezená kultivace byla srovnatelná mezi chemostat a fed-batch operace, vzhledem k tomu, že pro P-omezení, mírně se snížil výnos biomasy byl pozorován během fed-batch operace (Tabulky 2, 3), který je v rámci standardní odchylky pozorované u obou biomasy výnosy. Výtěžek uhlíku na substrát pro 3-HP byl 15,9% pro C-omezení, což je téměř totožné ve srovnání s hodnotou vypočtenou v chemostatovém systému (obr. 4a). 3-HP výnosů stanovených v této studii jsou dále v úzké dohody uhlíku výnos za substrát o 14%, získané v předchozí C-omezený fed-batch studium na pH 5,0 s S. cerevisiae navržen tak, aby použít β-alanin dráhy a 13% s S. cerevisiae s využitím malonyl-CoA reduktázy-závislé dráhy . Vyšší 3-HP uhlíku výnosy na substrát prostřednictvím β-alanin dráhy byly stanoveny ve studiích s Escherichia coli uvádí hodnotu 42%, fed-batch kultivace na glukóze .
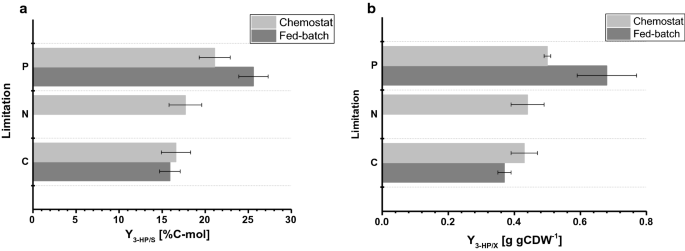
Porovnání pěstování parametrů v rámci různých omezení stanoví v malém měřítku chemostats a 1-L bench-top reaktorů ve fed-batch režim. výtěžek uhlíku 3-HP (%C-mol) a výtěžek b 3-HP na biomasu (g gCDW-1). Chyby odpovídají směrodatné odchylky odvozené z trojím vyhotovení pěstování
Pro P-omezující podmínky, nicméně naše studie odhalila značný nárůst 3-HP uhlíku výnos o 25,6% (Obr. 4a). Pozorované výtěžky 3-HP na biomase byly 0,38 a 0,65 g gCDW-1 pro podmínky omezující C A P. Pro C-omezení to potvrdilo hodnoty pozorované při nastavení chemostatu (obr. 4b). V důsledku zvýšení výtěžku 3-HP na substrát za podmínek omezujících P a mírného snížení výtěžku biomasy na substrát současně byl výtěžek 3-HP na biomasu významně zvýšen ve srovnání s experimentem s chemostatem. Na rozdíl od kultivací prováděných v chemostatech nebyla v kultivacích krmených šarží zjištěna žádná významná akumulace ethanolu nebo glycerolu. Uzavřená uhlíková bilance pro podmínky omezující C A P ukázala, že nebyla vytvořena žádná relevantní množství jiných (vedlejších)produktů (Tabulka 3). Tento rozdíl ve spektru vedlejších produktů může vysvětlit zvýšení výtěžku 3-HP na glukózu.
Ve srovnání se kultivace provádí v chemostats, specifické vychytávání glukózy sazby stanovena ve fed-batch kultur ukazují výrazně vyšší hodnoty 0,75 a 0,77 mmol gCDW−1 h−1 pro C – a P-omezující podmínky, respektive. Jak bylo uvedeno výše, nastavit bod pro exponenciální faktor krmiva profilu byla nastavena na 0,05 h−1, a lišily až o 0.059 h−1, což má za následek 30-45% vyšší specifická rychlost růstu ve fed-batch kultivace ve srovnání s chemostat experimenty s 0.04 h−1 (Tabulka 3), což způsobuje zvýšenou míru specifické absorpce glukózy. Nicméně, pro C-omezené fermentace, výtěžek biomasy na glukózu byl srovnatelný mezi chemostatem a kultivací krmených šarží. Podobně specifické 3-HP produkci sazby byly vyšší ve fed-batch kultivace, 0,24 mmol gCDW−1 h−1 a 0,38 mmol gCDW−1 h−1, resp. 3-HP výnosy na substrátu byly srovnatelné mezi chemostat a fed-batch kultivace, což naznačuje, výnosy, aby bylo robustní pěstování parametr, který je odolný vůči odchylky vyvolané menší výkyvy v pěstování set-up. Ve vzorcích odebraných z různých reaktorů pro podmínky omezující P nebyl zjištěn žádný zbytkový fosfát ani glukóza, což potvrzuje, že kultury mají být omezeny s ohledem na odpovídající omezení. Podobně pro C-omezení nebyla ve vzorcích zjištěna žádná zbytková glukóza.
zde prezentované vylepšení malého chemostatového systému, dříve vyvinutého Kleinem et al. , zahrnující mimo jiné zvýšení počtu reaktorů na 24 a snížení objemu kultivace na 6,5 ml. Tento zvýšený stupeň paralelizace je systém vhodný vysokokapacitní screening nástroj, jako různé srovnatelné malého rozsahu chemostat systémy pracují s vyšší pracovní objemy a nižší počet reaktorů: nepřetržité paralelní otřesený bioreaktoru (CosBios) systém používá šest, respektive osm paralelní kultivační nádoby s kulturou, objem 20-25 mL . Pro kontinuální kultivace modifikované jedno použití, míchá-nádrž bioreaktoru systému (bioreaktoru, 2mag AG) pracuje s osmi paralelních reaktorů v pracovním objemu 10 mL . Systém mini-chemostatu (MC) vyvinutý společností Bergenholm et al. obsahuje 16 paralelních reaktorů a vyžaduje pracovní objem 40 ml. Náš systém je proto dobře použitelný pro jednoduchý a nákladově efektivní screening mikrobiálního výkonu v nepřetržitém režimu. Mohlo by mít význam, nicméně, dále rozsah, systém sledování dalších parametrů, např. výfukových plynů, umožňují podrobnou analýzu uhlíku, rozdělení, což je zvláště důležité pro fyziologické napětí charakterizace.
Jako naše studie naznačila, N – a P-omezení být příznivé pro 3-HP formace v S. cerevisiae, budoucí experimenty by mohly zahrnovat další promítání růstu s N – a P-omezení aplikovaná s cílem najít optimální 3-HP produkci s každým příslušných omezení. Naše studie dále ukázala přenositelnost fyziologických parametrů z chemostatu na kultivace krmených šarží. To považuje za jediné možné, pokud žádné toxické nebo inhibiční sloučeniny se hromadí během fed-batch kultivace, od fyziologických parametrů by v průběhu času měnit a lišit od získané parametry v chemostats. Jako obecný přístup, chemostat experimenty mohou sloužit jako nástroj pro zkoumání vlivu potenciální toxické nebo inhibiční sloučeniny přidáním látky, aby se uživili. Protože všechny ostatní parametry jsou konstantní, lze přímo posoudit a vyhodnotit vliv sloučeniny a její koncentrace. V naší studii, tam byl žádné vedlejší produkt tvorby zjištěny ve fed-batch kultivace a pravděpodobně podobné reakce na stres byl vyvolán v důsledku slabé kyseliny, což umožnilo fyziologických podmínek v malém měřítku chemostats se podobají těm ve fed-batch podmínky.
stručně řečeno, tato studie ukázala, že koncept C – a P-omezující podmínky pro výrobu 3-HP byl sledován v románu parallelized chemostat pěstování systému, a může být úspěšně převedeny do 1-L bench-top bioreaktory působící ve fed-batch režim. Proto, fyziologické parametry získané v chemostatech mohou být použity pro návrh a hodnocení výkonu kultivací krmených šarží pomocí parametrů založených na výtěžnosti pro nastavení procesu.