
Buněčné Adheze Proteinů
Mobilní-buněčné adheze je selektivní proces, tak, že buňky dodržovat pouze na jiné buňky specifické typy. Tato selektivita byla poprvé prokázána v klasické studium vývoje embrya, která ukázala, že buňky z jedné tkáně (např. játra), konkrétně dodržovat buňky stejné tkáni, spíše než do buňky jiné tkáně (např. mozek). Takové selektivní buňky-buněčné adheze je zprostředkována transmembránové proteiny zvané buněčných adhezních molekul, která může být rozdělena do čtyř hlavních skupin: selectins, integrins, imunoglobulin (Ig) nadčeleď (tak pojmenovaný protože obsahují strukturní domény podobné imunoglobuliny), a cadherins (Tabulka 12.3). Adheze buněk zprostředkovaná selektiny, integriny a kadheriny vyžaduje Ca2+ nebo Mg2+, takže mnoho adhezivních interakcí mezi buňkami je závislých na Ca2+ nebo Mg2+.
tabulka 12.3
buněčné adhezní molekuly.
selektiny zprostředkovávají přechodné interakce mezi leukocyty a endoteliálními buňkami nebo krevními destičkami. Existují tři členové selektinu rodina: L-selektinu, která je vyjádřena na leukocyty; E-selektinu, který je exprimován na endoteliálních buňkách; a P-selektinu, který je exprimován na krevních destičkách. Jak bylo uvedeno výše v této kapitole, selektiny rozpoznávají sacharidy na povrchu buněk (viz obrázek 12.14). Jeden z jejich zásadní rolí je zahájit interakce mezi leukocyty a endoteliální buňky během migrace leukocytů z cirkulace do místa zánětu tkáně (Obrázek 12.62). Selektiny zprostředkovávají počáteční adhezi leukocytů k endoteliálním buňkám. Toto je následované tvorbou více stabilní adheze, v níž integrins na povrchu leukocytů vázat na mezibuněčné adhezivní molekuly (ICAMs), které jsou členy Ig nadčeleď exprimovány na povrchu endotelových buněk. Pevně připojené leukocyty jsou pak schopny proniknout stěnami kapilár a vstoupit do podkladové tkáně migrací mezi endoteliálními buňkami.

obrázek 12.62
adheze mezi leukocyty a endoteliálními buňkami. Leukocyty opouštějí cirkulaci v místech zánětu tkáně interakcí s endoteliálními buňkami kapilárních stěn. Prvním krokem v této interakci je vazba selektinů leukocytů (více…)
vazba ICAMs na integrins je příklad heterophilic interakce, ve které adhezní molekuly na povrchu jedné buňky (např. ICAM) rozpozná různé molekuly na povrchu jiné buňky (např. integrin). Ostatní členové nadčeledi ig zprostředkovávají homofilní interakce, ve kterých se adhezní molekula na povrchu jedné buňky váže na stejnou molekulu na povrchu jiné buňky. Taková homofilní vazba vede k selektivní adhezi mezi buňkami stejného typu. Například nervové buňky adhezní molekuly (N-CAMs) jsou členy Ig nadčeleď vyjádřil na nervové buňky, a homophilic vazba mezi N-CAMs přispívá k tvorbě selektivní asociace mezi nervových buněk během vývoje. Existuje více než 100 členů nadčeledi Ig, které zprostředkovávají různé interakce buněk a buněk.
čtvrtá skupina buněčných adhezních molekul, kadheriny, také vykazují homofilní vazebné specifika. Nejsou zapojeny pouze do selektivní adheze mezi embryonálními buňkami, ale jsou také primárně zodpovědné za tvorbu stabilních spojů mezi buňkami v tkáních. Například E-kadherin je exprimován na epiteliálních buňkách, takže homofilní interakce mezi E-kadheriny vedou k selektivní adhezi epiteliálních buněk k sobě navzájem. Je pozoruhodné, že ztráta E-cadherinu může vést k rozvoji nádorů vznikajících z epiteliálních buněk, ilustrující význam buňka-buňka v kontrole buněčného chování. Různé členové cadherin rodině, jako je N-cadherin (nervové cadherin) a P-cadherinu (placentární cadherin), zprostředkovávají selektivní adhezi jiné typy buněk.
bylo identifikováno asi dvacet různých klasických kadherinů, jako je E-cadherin. Kromě toho, odlišné podčeledi cadherins, tzv. protocadherins, jsou vyjádřeny v centrálním nervovém systému, kde se zdá, že hrát roli v adhezi mezi neurony, v synapsích. Zajímavé je, že různé neurony exprimují různé protocadheriny, což naznačuje, že protocadheriny mohou hrát roli při vytváření specifických spojení mezi neurony. Bylo identifikováno asi 50 lidských protocadherinových genů a ukázalo se, že jsou organizovány do tří genových shluků. Každý cluster obsahuje několik exonů kódující N-terminální extracelulární a transmembránové protocadherin domén, ale pouze jednu sadu tři exons kódující C-terminální cytoplazmatická doména (Obrázek 12.63). Zdá se tedy, že klastry genů protocadherin sestávají z proměnné oblasti, kódující více extracelulárních a transmembránových domén, spojených s konstantní oblastí kódující jednu cytoplazmatickou doménu. Tato organizace protocadherin geny nápadně podobá imunoglobulin a T-cell receptor geny (viz obrázek 5.42 5.43 a), ve kterém více variabilní oblasti exonů jsou spojeny do jedné konstanty regionu exon. V genech imunoglobulinu A T-buněčných receptorů k tomu dochází v důsledku přeskupení DNA, které generují rozmanitost v imunitním systému. Zbývá určit, zda je variabilní a konstantní oblasti protocadherins jsou spojeny v DNA nebo RNA úrovni (například tím, že alternativní sestřih) a do jaké míry přestavby z protocadherin geny by mohly přispět k vytvoření konkrétních synaptická spojení v mozku.
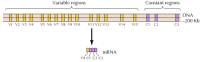
obrázek 12.63
organizace klastrů genů protocadherinu. Lidské protocadherinové geny jsou uspořádány do tří klastrů. V ilustrovaném klastru je 15 různých proměnných oblastí kódujících extracelulární a transmembránové domény spojeno s jednou konstantní oblastí, (více…)
na rozdíl od stabilní buňka-matrix uzlů popsaných v předchozím oddíle, mobilní-buněčné interakce zprostředkované selectins, integrins, a členové Ig nadčeleď jsou přechodné srůsty, ve kterém cytoskeletons sousední buňky nejsou vzájemně propojeny. Stabilní adhezní spoje zahrnující cytoskeletony sousedních buněk jsou místo toho zprostředkovány kadheriny. Jak je uvedeno v Kapitole 11, tyto mobilní-mobilní uzly jsou dvou typů: adherens junctions a desmozomy, ve kterém cadherins nebo související proteiny (desmogleins a desmocollins) jsou napojeny na aktin svazky a střední filamenta, respektive (Obrázek 12.64). Úloha kadherinů při propojení cytoskeletů sousedních buněk je tedy analogická s úlohou integrinů při vytváření stabilních spojů mezi buňkami a extracelulární matricí.

Obrázek 12.64
stabilní spojení mezi buňkami zprostředkované kadheriny. Homofilní interakce mezi kadheriny zprostředkovávají dva typy stabilních adhezí buněk a buněk. V adherens junctions jsou kadheriny spojeny se svazky aktinových vláken přes kateniny (viz obrázek (více…)