
Úvod
Buněčné nebo tkáňové obnovy a regenerace jsou dva maindevelopmental požadavky dospělých organismů. Oba processeshave jako výchozí bod populaci kmenových buněk, obvykle lokalizovaných na konkrétní prostředí, tzv. „niche“ , které providesthem potřebné signály pro udržení stemness vlastnosti, orto rozlišovat na požadovanou různých typů buněk (Obrázek 1). Proliferace a diferenciace kmenových buněk musí být koordinována smrt buněk, které je třeba vyměnit. Kromě toho procesyjako je migrace buněk, epigenetika a buněčná komunikace, jsou také nezbytné pro správnou obnovu buněk . Rychlé obnovenítkáně mohou být rozpoznány vyšší mitotickou aktivitou. Naopak,pomalé obnovy tkání obsahují méně mitózy, a nemusí být easilyrecognized z non-obnovují oblasti, které mohou také prezentovat somemitosis . Rozhodnutí o osudu kmenových buněk během proliferacepřímo ovlivnit obnovu tkáně a homeostázu. Proto pochopení regulačních mechanismů, které udržují vyváženérozdělení a diferenciace buněk je zásadní. Extracelulární signály (např. tkáňové mikroprostředí, intracelulární ROS, cytokiny) jakož i intracelulárních faktorů (např. epigenetické strojů, transcriptionfactors a poškození DNA reakce) jsou zodpovědné za nařízení, kmenových buněk, dělení.
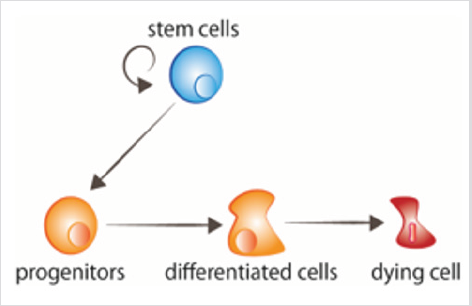
obrázek 1: obrat Buněk Kmenové buňky proliferují, givingrise se předci, že poté přijímat signály todifferentiate. Stárnoucí buňky přijímají signály k umírání.
Kmenové buňky ukazují tři možné varianty oddělení: asymetrické dělení, ve kterém jedna kmenová buňka a jedna committeddaughter buňky jsou vznikl; symetrické závazek, whichyields dva spáchal dcera buněk; a symetrické rozdělení,které přináší dvě dceřiné buňky, které udržují vlastnosti kmenových buněk Obrázek 2. I když by to mohlo být předpovídal, že asymmetricdivision je jediný mechanismus, který umožňuje údržbu ofa stabilní populace kmenových buněk, aktuální data z lineagetracingexperiments prokázala, že ve většině tkání, balancebetween kmenových buněk, proliferaci a generace differentiatedoffspring je dosaženo na úrovni celého kmenových buněk populace.Ztráta kmenových buněk v důsledku diferenciace nebo poškození buněk vyvolávásymetrické dělení k vyplnění této mezery . Po kmenové buněčné dělení, thecells, že sledovat proces diferenciace projít odlišnýetapy, které jsou definovány kombinací transkripce tj. kontrolovat činnost odpovídající repertoár genů, andallow jejich odhodlání a terminální diferenciace. Pro každý celllineage konečný produkt posloupnost rozhodnutí je specificdifferentiated buňka typu Obrázek 3a. Ve většině případů,buněčné identity – produkt normální diferenciace – je stablewithin tkání, a jeho údržba je zásadní pro normální tissuefunction. Taková stabilita je dosaženo prostřednictvím epigenetické regulace -např. histonů demetylací a acetylací – výsledky v heritablepatterns z tkáňově specifické genové exprese .
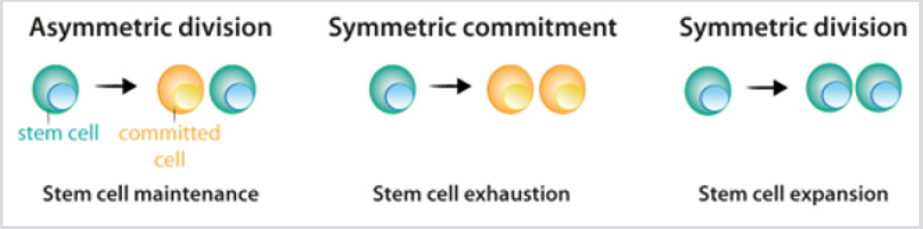
obrázek 2: Rozdělení vzor kmenových buněk.
a) během asymetrického dělení kmenové buňky vytvářejí jednu kmenovou buňku, která udržuje populaci kmenových buněk, a jednu buňku, která se zavazuje k diferenciaci.
b) během symetrického závazku vede dělení kmenových buněk ke vzniku dvou dceřiných buněk, které se zavázaly k diferenciaci.
c) během symetrického dělení vede dělení kmenových buněk ke vzniku dvou kmenových buněk. Jak je vysvětleno v textu, aktuální experimentálnídata naznačují, že tři způsoby dělení mohou nastat při zachování populace kmenových buněk.
může však dojít ke ztrátě buněčné identity. Buňky z imaginárního disku jsou totiž schopny transdeterminovat a získávat nový osud dospělých po transplantaci . V této situaci,extracelulární podněty se zdají přeprogramovat některé předchůdce ordifferentiated buňky získávají vlastnosti buď více stemstate nebo nové diferencované státu. Existují dva mechanismypodle kterých může buňka změnit svou identitu: dediferenciace a transdiferenciace. Dediferenciace se týká procesu pomocíkteré diferencovaná nebo oddaná buňka získává charakteristiky méně zralé buňky obrázek 3b. Nejdramatičtější partner dedifferentiation je in vitro konverze terminallydifferentiated buněk na pluripotentní buňky (vyvolané pluripotentstem buněk, iPSCs), zvýšená exprese omezený počet oftranscription faktory . Naproti tomu dochází k transdiferenciacekdyž diferencovaná buňka změní transkripční program apřevede se na jiný diferencovaný typ buňky. Proces můžedojde prostřednictvím zprostředkované krok dedifferentiation k aless zralé fázi před konverzí do nového differentiatedcell, nebo přímo, bez zprostředkovatelské fázi Obrázek 3c-d.Přímé přeměně fibroblastů do myoblastů, které ectopicexpression z MyoD je příkladem druhého procesu .
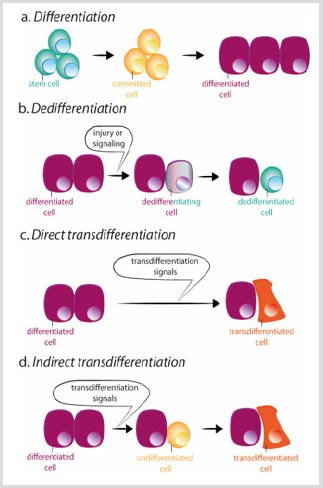
obrázek 3: Schémata diferenciace, dedifferentiation a trans-diferenciaci.
a) během normální diferenciace kmenové buňky vedou ke vzniku buněk, které se zase diferencují v různých typech buněk.
b) Dediferenciace spočívá v získání vlastností kmenových buněk diferencovanou buňkou. Může dojít k Trans diferenciacipřímým nebo nepřímým způsobem.
c) Při přímé trans-diferenciace, diferencované buňky získává transkripční program jiný typ buňky, usuallyclosely související, jako například exokrinní endokrinní buňky pankreatu, stává různé diferencované buňky.
d) během nepřímé trans diferenciace diferencovaná buňka před přijetím nového transkripčního programu druhého typu buňky.
Dedifferentiation a přeměňování také dojít ina přirozený způsob, jak v reakci na zranění, nebo tkáně .Dediferenciace se například vyskytuje přirozeně během limbregenerace u obojživelníků urodele. Po amputaci končetiny,buňky přilehlé k ránu dedifferentiate, které tvoří blastema thatconsists nediferencovaných buněk, které se množí a nakonec,redifferentiate do stejný typ buněk, aby vytvářet všechny komponenty ztracené končetiny. Přírodní přeměňování dochází nepřímo:za prvé, mobilní dedifferentiates, a pak přírodní developmentalprogram je aktivována, což umožňuje buňky diferencovat do nové linie. Tsonis a spolupracovníci popsali přirozenémechanismus transdiferenciace v mloku. Zjistili, že kdyčočky jsou odstraněny, pigmentované epiteliální buňky z dorzálního iristransdiferentiate a regenerují chybějící tkáň. K dosažení tohoto, pigmentové epitelu buněk, musí nejprve dedifferentiate andproliferate vytvořit nový objektiv buňky a pak diferencují na themature buněk čočky . V obou situacích – dedifferentiationand redifferentiation do stejný typ buněk nebo transdifferentiateto nový typ buněk – složité sítě signálních drah maycontrol na transkripční program získal tím, že každá buňka ve skvělý čas-bod.
hus, časoprostorová kontrola genové exprese je neustále vyžadována během homeostázy zvířat a během aregenerativního procesu. Nicméně, během regenerace buněk mustre-přizpůsobit nové situaci, která vyžaduje, aby moreprofound rozhodnutí na buněčné úrovni, často včetně procesů ofdedifferentiation a přeměňování, že během homeostasisare vzácné.
- Clevers H, Loh KM, Nusse R (2014) signalizace kmenových buněk. Integralprogram pro obnovu a regeneraci tkání WNT signalizace a řízení stemcell. Věda 346 (6205): 1-9.
- Leblond C P, Walker B (1956) Obnova buněčné populace 36 (2): 255-276.
- Roostaee A, Benoit YD, Boudjadi S, Beaulieu JF (2016) epigenetika v obnově střevních epiteliálních buněk. J Cell Physiol 231 (11): 2361-2367.
- Ito K, Ito K (2016) metabolismus a kontrola rozhodnutí o osudu buněk aobnovení buněk. Annu Rev Cell Dev Biol 32: 399-409.
- Simons BD, Clevers H (2011) strategie pro homeostatické kmenové buňky selfrenewalin dospělých tkání. Buňka 145 (6): 851-862.
- Yang Y, Akinci E, Dutton JR, Banga A, Slack JM W, et al. (2013) Fázespecifické přeprogramování jaterních buněk myších embryí na beta buněčný fenotyp. Mech Dev 130 (11-12): 602-612.
- Merrell AJ, Stanger BZ (2016) plasticita dospělých buněk in vivo: dediferenciace a transdiferenciace jsou zpět ve velkém stylu. Nat Rev MolCell Biol 17 (7): 413-425.
- Worley MI, Setiawan L, Hariharan IK (2012) Regenerace andtransdetermination v Drosophila imaginární disky. Annu Rev Genet 46: 289-310.
- Raff M (2003) plasticita dospělých kmenových buněk: fakt nebo artefakt? Annu Rev CellDev Biol 19: 1-22.
- Takahashi K, Yamanaka S (2006) indukce pluripotentních kmenových buněk z myších embryonálních a dospělých fibroblastových kultur definovanými faktory. Buňka 126 (4): 663-676.
- Tapscotr SJ, Davis RL, Thayer MJ, Cheng PF, Weintraub H, et al. (1988)MyoDi: a Nuclear Phosphoprotein Reqiring a Myc homology Region toConvert fibroblasty na Myoblasty. Věda 242 (4877): 405-411.
- Jopling C, Boue S, Izpisua Belmonte JC (2011) Dedifferentiation,přeměňování a přeprogramování: tři cesty k regeneraci.Nat Rev Mol Cell Biol 12 (2): 79-89.
- Tsonis PA, Madhavan M, Tancous EE, Del Rio-Tsonis K (2004)Del Rio-Tsonis, K. a mlok s pohled na regeneraci čoček. Int J Dev Biol 48 (8-9): 975-980.